
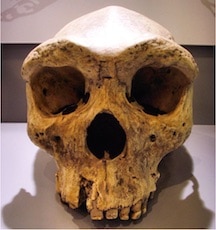
Image: Gerbil/Wikicommons
Early Hominin and the Auditory System
Studying the sensory mechanism fossil remains of past hominin (members of the family of humans as described herein) life forms has offered some interesting insights into ancient ears, based on their anatomical structures. This is especially true for the sense of hearing.
How this occurs relates to the physical properties of the fossil physical structure. For the hearing mechanism, the anatomy of the outer and middle ear assists in suggesting their auditory capabilities. Of all the senses, the ear (audition) is especially amenable to study because audition is strongly related to physical structures that are best preserved in fossil specimens.
A recent publication on “Early hominin auditory capacities”{{1}}[[1]]Quam,R. Martinez, I., Rosa, M., Bonmati, A., Lorenzo, C., de Ruiter, D., Moggi-Cecchi, J., Valverde, M. Jarabo, P., Menter, C., Thackeray, J., and Arsuaga, J., Early hominin auditory capacities, Science Advances, Vol. 1., no. 8, September, 2015[[1]], reported on their investigation into measurements of the taxonomy of Australopithecus africanus (A. africanus) and Paranthropus robustus (P. robustus) to estimate their auditory capacities. The data in this post is extracted from that article.
The Australopithecus africanus lived about 3.3 to 2.5 million years ago in South and East Africa, and the Paranthropus robustus, also from South Africa, about 2.0 to 1.4 million years ago as shown in Figure 1.
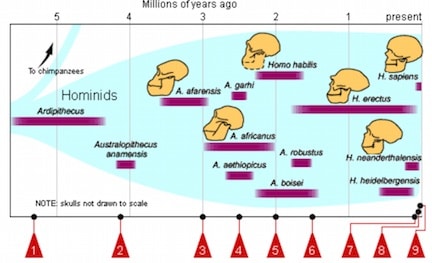
Figure 1. Time line of hominid progression. fhs-bio-wiki.pbworks.com
Compared to Today’s Humans
As compared to current humans, these early hominin taxa had slightly shorter and wider external auditory canals, smaller tympanic membranes, and a lower ossicular lever action ratio. Additionally, they appeared to have a smaller size stapes footplate, as calculated from the annular ring, which also contributed to the speculated lower lever action ratio. Because of these measurement differences, it was suggested that these dimensions may have facilitated an increased emphasis on short-range vocal communication.
Previous studies comparing later hominin development (Middle Pleistocene) had shown similar auditory capacities as living humans. However, an emerging field of inquiry studying the sensory ecology and communication in early hominins suggested new insights into their physical adaptations.
For example, might the anatomical physical dimensions provide information relative to hearing range and spectra, and from this, theories as to what frequency range their vocalizations consisted of? Of major significance in all these reports, is that the sample size is very small, in some cases consisting of only a single skull.
Measurement Tools Used
CT scans and virtual reconstructions of the outer and middle ear were used for measurement purposes, along with direct measurements on the original fossils when the outer and middle ear was exposed and accessible. Ossicles were scaled from digital images following removal from the tympanic cavity. In some cases, data was taken from previous publications. In some cases, the dimensions were estimated. Any soft tissue variables used values of modern humans. Middle ear modeling capacities were based on a modified Rosowski model{{2}}[[2]]J. Rosowski, in Auditory Computation, H. Hawkins, T. McMullen, A. Popper, R. Fay, Eds. Springer, New York, 1996), pp. 15–61[[2]].
Overall Findings
Those structures that had the greatest influence on the model results were from the outer ear and the auditory ossicles. Middle ear spaces (tympanic cavity, the air-celled mastoid, and atrium) had a lesser influence on the results.
Compared with humans, A. africanus showed significant differences in three of six skeletal variables (tympanic cavity volume, ear canal length, and stapes footplate area). P. robustus produced significant differences from living humans on ear canal length, ear canal cross sectional area, and tympanic membrane area.
A comparison between the two early hominin taxa, A. africanus and P. robustus was limited, having just four variables with sample size of n>3. The differences were the tympanic membrane area and stapes footplate, with both larger in P. robustus.
The differences toward modern humans are that the early hominin had slightly shorter and wider ear canals, smaller tympanic membranes, a lower malleus/incus lever ratio, and a smaller stapes footplate.
A model calculation of the sound power transmission values from 0 to 5000 Hz is provided in Figure 2. This model is reflected in the frequency range (bandwidth) that concentrates more than 90% of the sound power that reaches the inner ear.
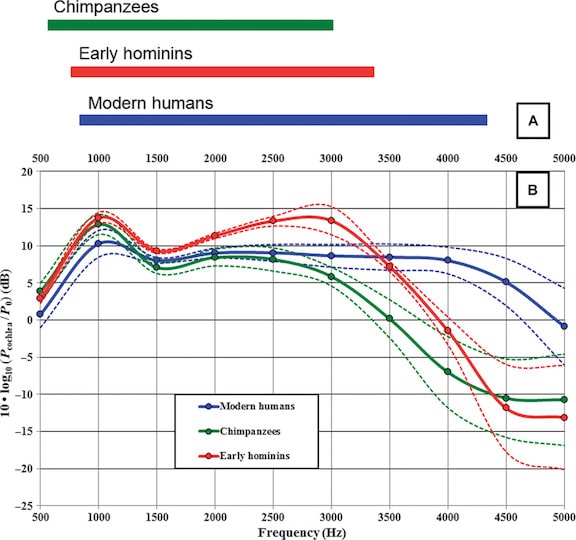
Figure 2. Model results for chimpanzees, modern humans, and early hominins from 0.5 to 5.0 kHz. (A) The occupied band is similar in chimpanzees and early hominins, but is shifted toward slightly higher frequencies in the latter. Modern humans show a widened occupied band that is further extended toward higher frequencies. (B) The sound power transmission curves correspond to decibels at the entrance to the cochlea relative to P0 = 10−18 W for an incident plane wave intensity of 10−12 W/m2. The mean value ± 1.0 (Quam, et. al., 2015).
Based on this model, early hominins are more sensitive than humans between 1.5 and 3.0 kHz. (This post was not interested in comparisons to chimpanzees). Later hominins (middle Pleistocene Atapuerca) have been shown to have sound power transmission close to that of modern humans.
Resonance characteristics of the ear canal can be approximated as four times the length of the tube (Lec x 4). Based on ear canal lengths reported of 28.6 and 29.7mm, this would place the ear canal resonant frequencies at 3.0 and 2.9 kHz respectively for the A. africanus and P. robustus and is directly related to both cutoff frequencies.*
Ear canal width in both hominins was measured slightly larger than humans, the tympanic membrane area was close to that of humans, but the ossicular level ratio of P. robustus was somewhat higher.
Potential Impact on Vocalizations
The wider occupied power response range of modern humans, especially in the extended upward frequencies, is speculated to facilitate complex short-range vocal communication. This may be true for high-frequency consonant sounds, which is one of the salient features of human spoken language versus animal communications.
In contrast, suggestions that early hominins’ communication may have been largely restricted to a vowel-based system have been advanced, hinting at the reason for the sound power transmission changes in the 2000 to 3000 Hz region.
*Editor’s note: These ear canal lengths are reported longer than the approximate 25 mm lengths in measured published data on humans, so the report that they are shorter in the hominins measured could be a result of the small sample sizes.


